
遺伝子組み換え微生物を用いた高分子原料の発酵生産
高分子原料など工業原料として利用可能な化合物を遺伝子組み換え微生物を用いて発酵生産するための技術開発に取り組んでいます。自然界いる微生物をそのまま利用するだけでは発酵生産できる高分子原料の種類や量は限られてしまいます。そこで、遺伝子を人工的に操作した微生物を用いることで、本来生産できない高分子原料を作らせたり生産量を高めたりすることができます。発酵生産して得られる化合物はバイオマス由来なのでカーボンニュートラルの点で環境に優しい素材といえます。また、微生物を用いるため、反応は立体・位置選択性に優れており、グリーンケミストリーの考え方に即した低環境負荷型のモノづくりが可能です。
これまでに発酵生産した化合物の一覧を図2.1に示します。イタコン酸は大腸菌(J. Gen. Appl. Microbiol., 60:191 (2014), J. Biosci. Bioeng., 119:548 (2015), Bioegneered, 6:303 (2015))とシアノバクテリア(J. Biotechnol., 195:43 (2015))、D-乳酸は大腸菌(J. Biosci. Bioeng., 127:441 (2019))と乳酸菌(Curr. Microbiol., 76:1186 (2019))、グリオキシル酸は大腸菌(Fermentation, 9:534 (2023))、グリセリン酸は大腸菌(J. Biosci. Bioeng., 135:375 (2023))、1,2-プロパンジオールは大腸菌(Curr. Microbiol., 77:3704 (2020))と乳酸菌(AMB Exp., 11:117 (2021))、1,3-プロパンジオールは大腸菌(Lett. Appl. Microbiol., 72:68 (2021))をそれぞれ遺伝子組み換えして発酵生産しています。

図2.1 これまでに発酵生産した化合物
研究の流れを図2.2に示します。代謝工学と発酵工学を組み合わせて目的化合物を発酵生産させる手法を取ります。
生産宿主となる微生物に対して、目的化合物の生産に関わる遺伝子を導入し発現させるとともに、副成物の生成など不要な反応に関わる遺伝子を破壊することで、基質(バイオマス)から効率的に目的生産物を生産する微生物を作出します。このように、目的化合物の生産のために人為的に代謝経路をデザインする研究を「代謝工学」と呼びます。研究室で対象とする生産宿主は、主に大腸菌、乳酸菌、シアノバクテリアなどの真正細菌(バクテリア)です。遺伝子組み換えを要するため、遺伝子発現のための技術(発現プロモーター、ベクター、形質転換法など)と遺伝子破壊のための技術(ゲノム上遺伝子の相同組換えなど)が確立された微生物を用います。
次に、作出した生産菌の発酵生産のパフォーマンスを最大限高めることで、より効率的に目的化合物の生産が可能となります。このように、目的化合物の生産のために微生物の培養条件(培地組成、pH、温度、溶存酸素濃度など)や培養方法(pH-stat流加培養、セルリサイクル培養など)などを工学的に改良する研究を「発酵工学」と呼びます。

図2.2 研究の流れ
研究の流れを図2.2に示します。代謝工学と発酵工学を組み合わせて目的化合物を発酵生産させる手法を取ります。
生産宿主となる微生物に対して、目的化合物の生産に関わる遺伝子を導入し発現させるとともに、副成物の生成など不要な反応に関わる遺伝子を破壊することで、基質(バイオマス)から効率的に目的生産物を生産する微生物を作出します。このように、目的化合物の生産のために人為的に代謝経路をデザインする研究を「代謝工学」と呼びます。研究室で対象とする生産宿主は、主に大腸菌、乳酸菌、シアノバクテリアなどの真正細菌(バクテリア)です。遺伝子組み換えを要するため、遺伝子発現のための技術(発現プロモーター、ベクター、形質転換法など)と遺伝子破壊のための技術(ゲノム上遺伝子の相同組換えなど)が確立された微生物を用います。
次に、作出した生産菌の発酵生産のパフォーマンスを最大限高めることで、より効率的に目的化合物の生産が可能となります。このように、目的化合物の生産のために微生物の培養条件(培地組成、pH、温度、溶存酸素濃度など)や培養方法(pH-stat流加培養、セルリサイクル培養など)などを工学的に改良する研究を「発酵工学」と呼びます。
ここからは、これまで実施してきた具体的な研究内容について、シアノバクテリアを用いたイタコン酸生産、大腸菌を用いたD-乳酸生産、乳酸菌を用いた1,2-プロパンジール生産を例に紹介します。
1. シアノバクテリアを用いたイタコン酸生産(J. Biotechnol., 195:43 (2015))
シアノバクテリアは光合成によりCO2から様々な化合物を生産することができます。本研究では、シアノバクテリアSynechocystis sp. PCC6803を用いて植物バイオマスを介さずCO2からダイレクトにイタコン酸を発酵生産するための技術開発に取り組みました。イタコン酸はTCA回路の中間代謝物であるcis-アコニット酸が酵素的に脱炭酸されて生成します。その反応を触媒する糸状菌Aspergillus terreus由来cis-アコニット酸脱炭酸酵素をコードする遺伝子cadを発現ベクターpJAK12を用いてSynechocystis sp. PCC6803に導入・発現させました。遺伝子組み換えシアノバクテリアをLEDライトを当てながら空気または5%CO2を通気しながら培養しました(図2.3)。その結果、それぞれ0.9 mg/Lまたは14.5 mg/Lのイタコン酸を発酵生産することができました(図2.4)。生産性を高めることで、工業的にCO2からイタコン酸を生産できるようになると期待されます。

図2.3 シアノバクテリアを用いたイタコン酸生産

図2.4 遺伝子組み換えシアノバクテリアによるCO2からのイタコン酸生産(青, 菌体; 白, イタコン酸)
(左)空気通気、(右)5%CO2通気
2. 大腸菌を用いたD-乳酸生産(J. Biosci. Bioeng., 127:441 (2019))
大腸菌は増殖性が良く遺伝子組み換えも容易なため、様々な化合物の生産宿主として良く使用されます。本研究では、大腸菌Escherichia coli BW25113を用いてセロビオースからD-乳酸を効率よく発酵生産するための技術開発に取り組みました。セロビオースはセルロースの酵素分解で生じる二糖です。D-乳酸の生産性を高めるために大腸菌ゲノム上のピルビン酸-ギ酸リアーゼ遺伝子pflAを破壊しました。次に、放線菌Thermobifida fusca YX由来のβ-グルコシダーゼ遺伝子bglCを発現ベクターpGV3を用いて、pflA破壊大腸菌の細胞表層に発現させました(図2.5)。pGV3は先の研究で開発した緑膿菌Pseudomonas aeruginosa由来リポタンパク質遺伝子oprI'を持つベクターで、oprI'の下流に酵素の遺伝子を融合させることで大腸菌の細胞表層に酵素を提示させることができます(J. Biosci. Bioeng., 119:548 (2015))。β-グルコシダーゼはセロビオースをグルコースに加水分解する酵素ですが、大腸菌の細胞表層にβ-グルコシダーゼを提示させセロビースをグルコースに変換して大腸菌が直接資化できるようにしました。ジャーファーメンターを用いて10 g/Lのセロビオースを含む培地で遺伝子組み換え大腸菌を嫌気条件下で回分培養しました。その結果、培養84時間後に5.2 g/LのD-乳酸を発酵生産することができました。次に、連続かつ効率的に発酵生産するために、菌体リサイクル発酵のシステムを開発しました。培養液上清と菌体を分離するための中空糸膜モジュールを用いて、培養液上清を回収し分離した菌体を再度培養液に戻すことで、大腸菌を高密度培養しながらD-乳酸を連続発酵生産するシステムとしました(図2.6)。菌体リサイクル発酵の結果、培養液中のD-乳酸濃度を4.3~5.0 g/Lに一定に保ちながら0.22–0.25 g/L/hの生産性でD-乳酸を連続発酵生産することに成功しました(図2.7)。連続発酵の生産性は回分培養の場合と比べて3倍以上に向上しました。
図2.5 大腸菌へのβ-グルコシダーゼの細胞表層発現
(左)実験概要 (中央)プラスミド作製 (右)β-グルコシダーゼ発現の確認
図2.6 菌体リサイクル発酵
(左)装置概要 (右)装置運転時のモデル



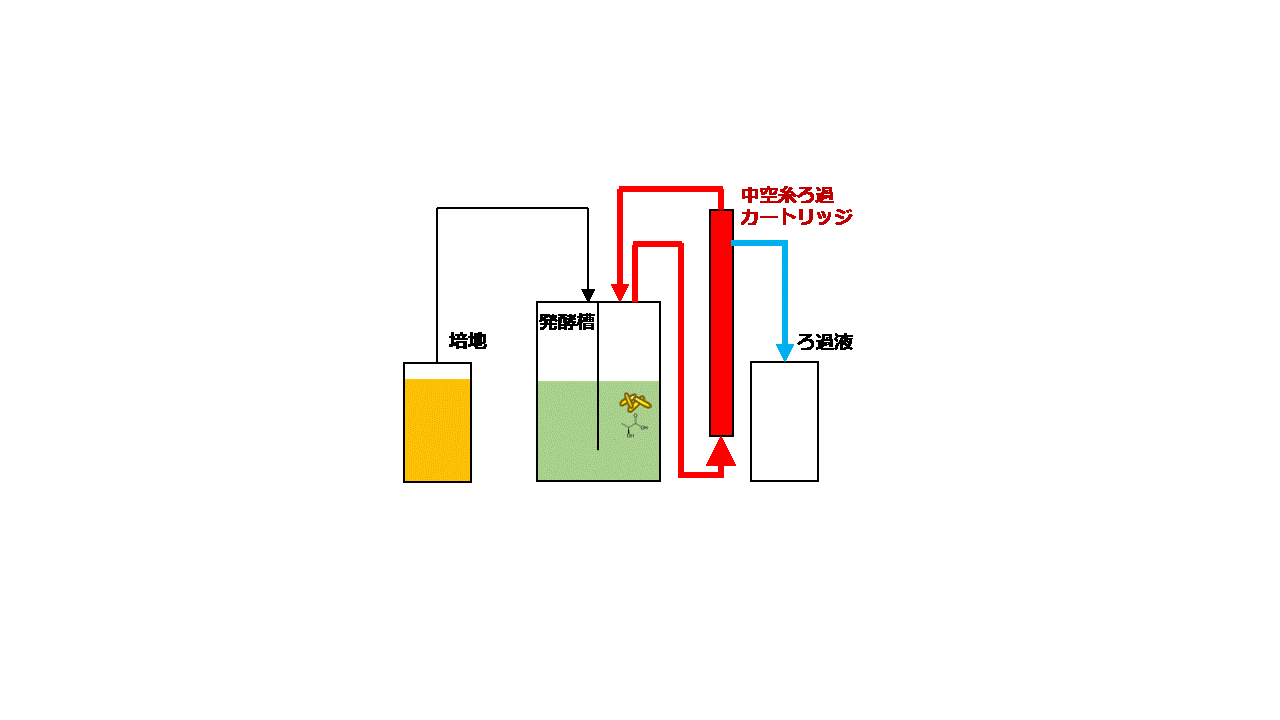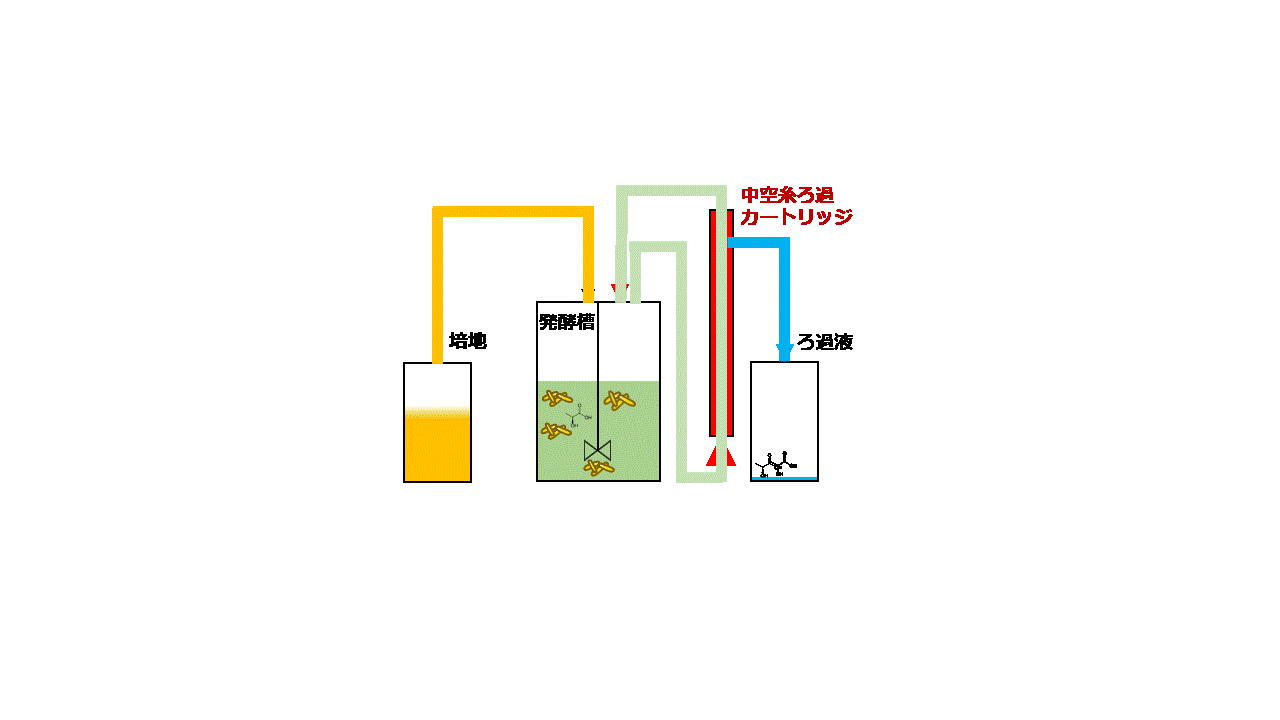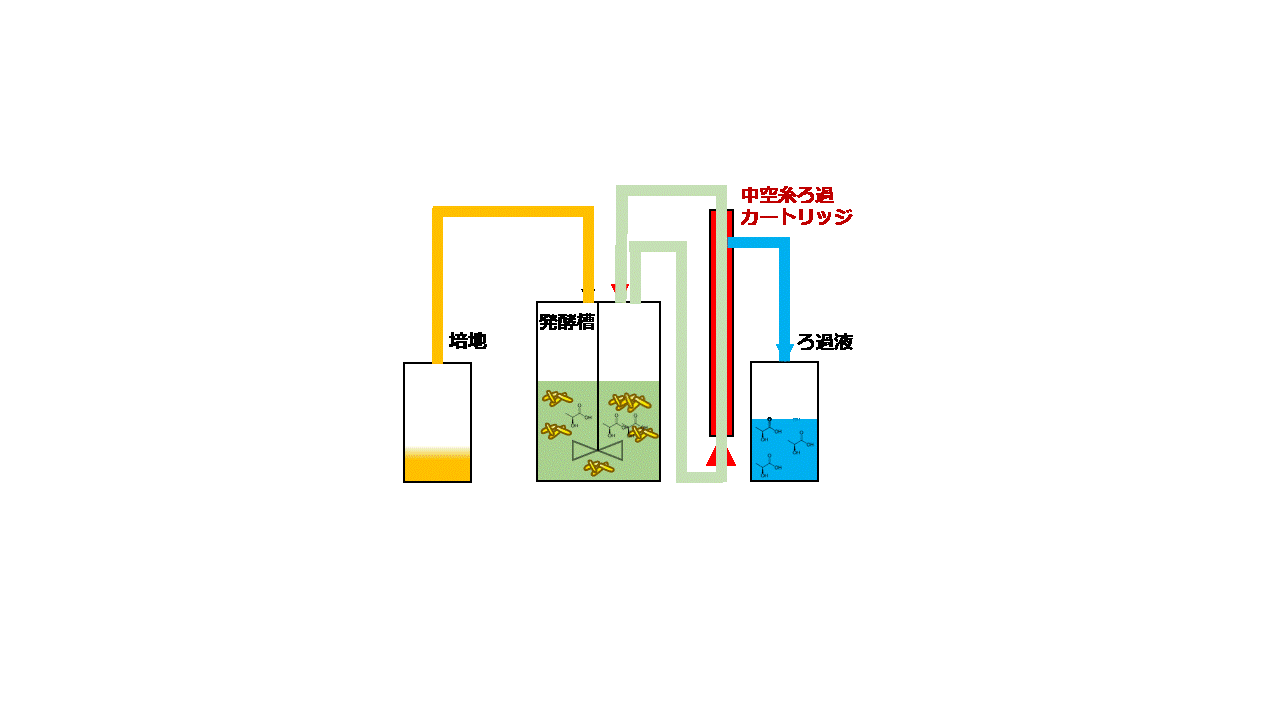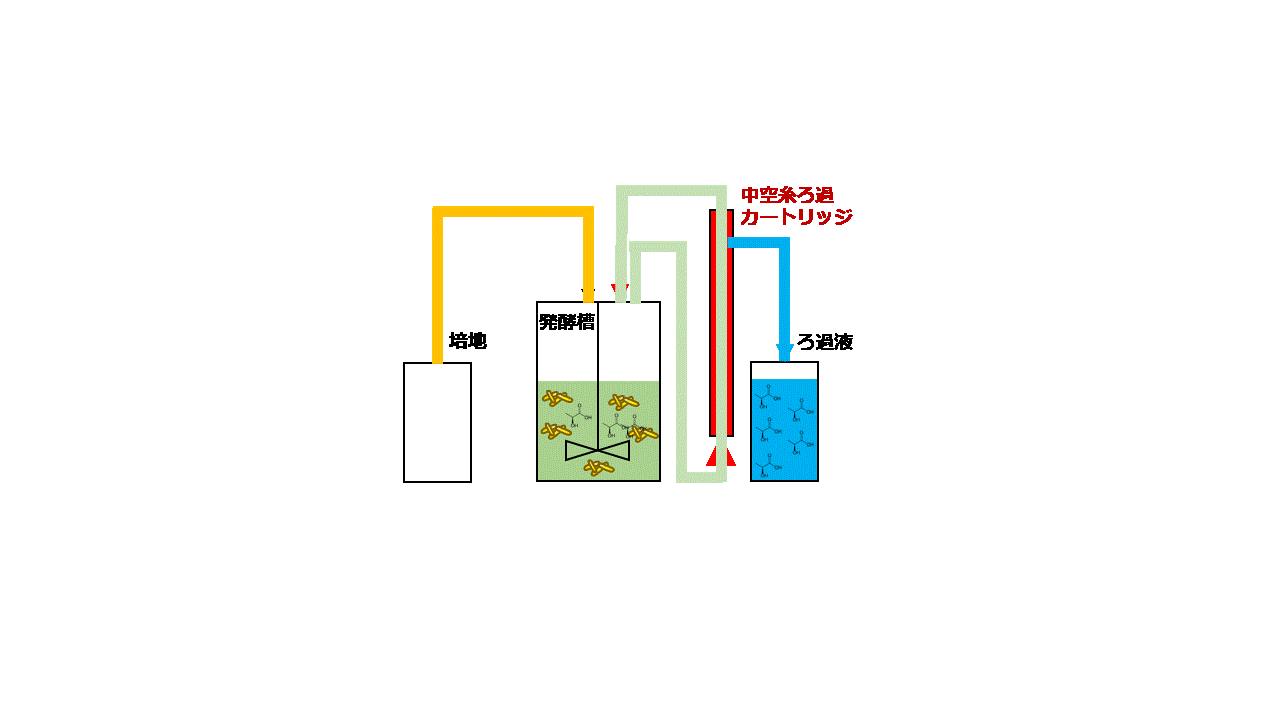

図2.7 遺伝子組み換え大腸菌によるセロビオースからのD-乳酸生産(菌体リサイクル発酵)
3. 乳酸菌を用いた1,2-プロパンジオール生産(AMB Exp., 11:117 (2021))
1,2-プロパンジオールの生産性を高めるために、1,2-プロパンジオールの前駆物質である乳酸を多量に生産する乳酸菌を用いて発酵生産を試みました。ルーメン細菌Megasphaera elsdenii由来プロピオン酸-CoAトランスフェラーゼ遺伝子pct、サルモネラ菌Salmonella enterica由来アルデヒドデヒドロゲナーゼ遺伝子pduP、大腸菌E. coli由来ラクトアルデヒドレダクターゼ遺伝子yahKを乳酸菌において導入・発現させ、グルコースから乳酸を経て1,2-プロパンジールを生産するための代謝経路をデザインしました(図2.8)。また、L-乳酸およびD-乳酸から(S)-1,2-プロパンジオールおよび(R)-1,2-プロパンジオールをそれぞれ作り分けることができます。そこで、L-乳酸およびD-乳酸をそれぞれ生産する乳酸菌Lactococcus lactis NZ9000およびL. lactis AH1において発現ベクターpNZ9000を用いてpct、pduP、yahKを導入・発現させました。L. lactis AH1は先の研究で開発したD-乳酸を生産し遺伝子組み換えが容易な乳酸菌です(Curr. Microbiol., 76:1186 (2019))。1%グルコースを炭素源として遺伝子組み換え乳酸菌をジャーファーメンターを用いて培養した結果、培養72時間後にそれぞれ0.69 g/Lの(S)-1,2-プロパンジオールと0.50 g/Lの(R)-1,2-プロパンジオールを生産することに成功しました。得られた乳酸の光学純度はそれぞれ94.4%eeと78.0%eeであり、(S)-1,2-プロパンジオールおよび(R)-1,2-プロパンジオールを作り分けられることが示されました。PduPとYahKはそれぞれ補酵素としてNADHとNADPHを要求します。そこで更なる生産性向上のために、炭素源としてそれらの各供給源となるマンニトールとグルコン酸を用いることにしました。5%マンニトールと5%グルコン酸を炭素源として遺伝子組み換え乳酸菌をジャーファーメンターを用いて培養した結果、培養96時間後に1.88 g/Lの(S)-1,2-プロパンジオールを生産することができました(図2.9)。

図2.8 遺伝子組み換え乳酸菌の代謝経路

図2.9 遺伝子組み換え乳酸菌によるマンニトールとグルコン酸からの1,2-プロパンジール生産